Archaeopteryx
2007 Schools Wikipedia Selection. Related subjects: Birds
| iArchaeopteryx |
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 A model of Archaeopteryx lithographica
on display at the Oxford University Museum |
||||||||||||||
| Scientific classification | ||||||||||||||
|
||||||||||||||
|
|
||||||||||||||
| Archaeopteryx lithographica Meyer, 1861 |
||||||||||||||
|
|
||||||||||||||
|
see text |
Archaeopteryx (from Ancient Greek αρχαιος archaios meaning 'ancient' and πτερυξ pteryx meaning 'feather' or 'wing'; pronounced /ɑː(ɹ)kiːˈɒptəˌɹɪks/), from the late Jurassic Period ( Kimmeridgian stage, 155-150 million years ago) of what is now Germany, is the earliest and most primitive known avian. Archaeopteryx was similar in size and shape to a magpie, with broad, rounded wings and a long tail. Its feathers resembled those of modern birds but Archaeopteryx was rather different from any bird known today, in that it had jaws lined with sharp teeth, three 'fingers' ending in curved claws and a long bony tail. In 1862, the description of the first intact specimen of Archaeopteryx, just two years after Charles Darwin published The Origin of Species, set off a firestorm of debate about evolution and the role of transitional fossils that endures to this day.
Archaeopteryx and the origins of birds
In the 1970s, John Ostrom argued that the birds evolved from theropod dinosaurs (see Dinosaur-bird connection). Archaeopteryx provides a critical piece of this argument, as it preserves a number of avian features (a wishbone, flight feathers, wings, a partially reversed first toe) and a number of dinosaur and theropod features (for instance, a long ascending process of the astragalus, interdental plates, an obturator process of the ischium, and long chevrons in the tail). In particular, Ostrom found that Archaeopteryx was remarkably similar to the theropod family Dromaeosauridae. Further research on dinosaurs from the Gobi Desert and China has since provided more evidence of a link between Archaeopteryx and the dinosaurs, such as Chinese feathered dinosaurs.
Archaeopteryx is probably close to the ancestry of modern birds - it shows most of the features one would expect in an ancestral bird - but it may not be the direct ancestor of living birds, and it is arguable how much divergence was already present in the early birds at its time.
Plumage
Archaeopteryx specimens were most notable for their well-developed remiges. These are markedly asymmetrical and show the structure of flight feathers of modern birds, with vanes given stability by a barb- barbule-barbicel arrangement. The tail feathers are less asymmetrical, again in line with the situation in modern birds, and also have firm vanes. The thumb did not bear a separately movable tuft of stiff feathers ( alula) yet.
Body plumage is less well documented, and only properly researched in the well-preserved Berlin specimen. Thus, as more than one species seems to be involved, the following does not necessarily hold true for all of them. In the Berlin specimen, there are "trousers" of well-developed feathers on the legs; some of these feathers seem to have a basic contour feather structure but are somewhat decomposed (i.e., lack barbicels as in ratites: Christiansen & Bonde, 2004), but at least in part they are firm and thus capable of supporting flight (Longrich, 2006).
There was a patch of pennaceous feathers running along the back which was quite similar to the contour feathers of the body plumage of modern birds in being symmetrical and firm (though not as stiff as the flight-related feathers). Apart from that, the feather traces in the Berlin specimen are limited to a sort of "proto- down" not dissimilar to that found in the dinosaur Sinosauropteryx, being decomposed and fluffy, and possibly even appeared more like fur than like feathers in life (though not in their microscopic structure). These occur on the remainder of the body, as far as such structures are both preserved and not obliterated by preparation, and the lower neck (Christiansen & Bonde, 2004).
On the other hand, there is no indication of feathering on the upper neck and head; while these may conceivably have been nude as in many closely related feathered dinosaurs for which good specimens are available, this may still be an artifact of preservation: it appears that most Archaeopteryx specimens became embedded in anoxic sediment after drifting some time on their back in the sea - the head and neck and the tail are generally bent downwards which suggests that the specimens had just started to rot when they were embedded, with tendons and muscle relaxing so that the characteristic shape of the fossil specimens was achieved. This would mean that the skin was already softened and loose (further evidence is provided by the fact that in some specimens, the flight feathers were starting to detach at the point of embedding in the sediment), and in specimens moving along the ground in shallow water, this would cause the head and upper neck, but not the more firmly attached tail feathers to slough off (Elżanowski, 2002).
It must be mentioned that the feather, the initial specimen described, does not agree too well with the flight-related feathers of Archaeopteryx. It certainly is a remix of a contemporary species, but its size and proportions indicate that it probably belongs to an as of yet undiscovered species of primitive bird or possibly bird-like dinosaur. As the feather was the original type specimen, this has created quite some nomenclatorial confusion.
Flight ability
The flight feathers of Archaeopteryx were highly asymmetrical, as in the wings of modern birds, and the tail feathers are rather broad. This implies that the wings and tail were used for lift generation, but it is unclear whether Archaeopteryx was simply a glider, or capable of flapping flight. The lack of a bony breastbone suggests that Archaeopteryx was not a very strong flier, but flight muscles might have attached to the thick, boomerang-shaped wishbone, the platelike coracoids, or perhaps to a cartilagenous sternum. The sideways orientation of the glenoid (shoulder) joint between scapula, coracoid and humerus - instead of the dorsally angled arrangement found in modern birds - suggests that Archaeopteryx was unable to lift its wings above its back, a requirement for the upstroke found in modern flapping flight. Thus, it seems likely that Archaeopteryx was indeed unable to use flapping flight as modern birds do, but it may well have utilized a downstroke-only flap-assisted gliding technique (Senter, 2006).
Archaeopteryx wings were relatively large, which would have resulted in a low stall speed and reduced turning radius. The short and rounded shape of the wings would have increased drag, but could also have improved Archaeopteryx' ability to fly through cluttered environments such as trees and brush (similar wing shapes are seen in birds which fly through trees and brush, such as crows and pheasants). The presence of "hind wings", asymmetrical flight feathers stemming from the legs similar to those seen in dromaeosaurids such as Microraptor, would also have added to the aerial mobility of Archaeopteryx. The first detailed study of the hind wings by Longrich (2006) suggested that the structures formed up to 12% of the total airfoil. Considering that it is not certain to what extent such feathers capable of supporting flight were present on the legs, this would have reduced stall speed by up to 6% and turning radius by up to 12%, in addition to the stall and turning radius reduction provided by the primary wing and tail feathers.
In 2004, scientists analyzing a detailed CT scan of Archaeopteryx's braincase, concluded that its brain was significantly larger than that of most dinosaurs, indicating that it possessed the brain size necessary for flying. The overall brain anatomy was reconstructed using the scan. The reconstruction showed that the regions associated with vision took up nearly one-third of the brain. Other well-developed areas involved hearing and muscle coordination (Winter, 2004). The skull scan also revealed the structure of the inner ear. The structure more closely resembles that of modern birds than the inner ear of reptiles. These characteristics taken together suggest that Archaeopteryx had the keen sense of hearing, balance, spatial perception and coordination needed to fly (Alnso et al., 2004).
Archaeopteryx continues to play an important part in scientific debates about the origin and evolution of birds. Some scientists see Archaeopteryx as a semi-arboreal climbing animal, following the idea that birds evolved from tree-dwelling gliders (the "trees down" hypothesis for the evolution of flight proposed by O.C. Marsh). Other scientists see Archaeopteryx as running quickly along the ground, supporting the idea that birds evolved flight by running (the "ground up" hypothesis proposed by Samuel Wendell Williston). Still others suggest that Archaeopteryx might have been at home both in the trees and on the ground, like modern crows, and this latter view is what today is considered best-supported by morphological characters. Altogether, it appears that it was a species which was neither particularly specialized for running on the ground, nor for perching. Considering the current knowledge of flight-related morphology, a scenario as outlined by Elżanowski (2002), namely that Archaeopteryx used its wings mainly to escape predators by glides punctuated with shallow downstrokes to reach successively higher perches, and alternatively to cover longer distances by (mainly) gliding down from cliffs or treetops, appears quite reasonable.
Given that it is now well established that several lineages of theropods evolved feathers and flight independently, the question of how precisely the ancestors of Archaeopteryx became able to fly has lost dramatically in importance for the time being. Since it is quite likely that this species belongs to a lineage of birds unrelated to the Neornithes (the Jurassic ancestor of which remains unknown), how exactly flying ability was gained in Archaeopteryx may be a moot point, having little bearing on how this happened in the ancestors of modern birds.
Taxonomy
The relationships of the specimens are problematic. Most specimens have been given their own species at one point or another. The Berlin specimen has been referred to Archaeornis siemensii, the Eichstätt specimen to Jurapteryx recurva, the Munich specimen to Archaeopteryx bavarica and the Solnhofen specimen was referred to Wellnhoferia grandis.
Recently, it has been argued that all the specimens belong to the same species ( New Scientist, 17 April 2004, p.17). However, significant differences exist among the specimens. In particular, the Munich, Eichstätt, Solnhofen and Thermopolis specimens differ from the London, Berlin, and Haarlem specimens in being smaller or much larger, having different finger proportions, having more slender snouts, lined with forward-pointing teeth and possible presence of a sternum. These differences are as large as or larger than the differences seen today between adults of different bird species. However, it is also possible that these differences could be explained by different ages of the living birds.
Fossils
Over the years, ten specimens of Archaeopteryx have been found. All of the fossils come from the limestone deposits near Solnhofen, Germany.
- The feather: Discovered in 1860 near Solnhofen, Germany, and described in 1861 by Hermann von Meyer. Currently located at the Humbolt Museum für Naturkunde in Berlin. This is generally referred to Archaeopteryx and was the initial holotype, but whether it actually is a feather of this species or another, as yet undiscovered, proto-bird is unknown. There are some indications it is indeed not from the same animal as most of the skeletons (the "typical" A. lithographica) (Griffiths, 1996).
 The London Archaeopteryx, 1863, detail, note the feathers
The London Archaeopteryx, 1863, detail, note the feathers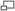
- London Specimen (BMNH 37001): Discovered in 1861 near Langenaltheim, Germany and described in 1863 by Richard Owen as Archaeopteryx macrura, assuming it did not belong to the same species as the feather. Currently located at the British Museum of Natural History in London, it is missing its head. In a subsequent edition of his Origin of Species (chap. 10, pp.335-336), Charles Darwin acclaimed Owen's discovery as linking lizard-like reptiles with modern birds.
 The Berlin Archaeopteryx, 1881
The Berlin Archaeopteryx, 1881 - Berlin Specimen (HMN 1880): Discovered in 1876 or 1877 on the Blumenberg near Eichstätt, Germany, by Jakob Niemeyer. He exchanged this precious fossil for a cow, with Johann Dörr. It was described in 1884 by Wilhelm Dames. Currently Located at the Humbolt Museum für Naturkunde, it is the best specimen, and the first with a complete head. Once classified as a new species, A. siemensii, but a recent evaluation supports the A. siemensii species definition [Elzanowski, 2002].
- Maxberg Specimen (S5): Discovered in 1956 or 1958 near Langenaltheim and described in 1959 by Heller. Currently missing, though it was once exhibited at the Maxberg Museum in Solnhofen. It belonged to Eduard Opitsch, who loaned it to the museum. After his death in 1991, the specimen was discovered to be missing and may have been stolen or sold. It is composed of a torso.
- Haarlem Specimen (TM 6428, also known as the Teyler Specimen): Discovered in 1855 near Riedenburg, Germany and described as a Pterodactylus crassipes in 1875 by von Meyer, it was reclassified in 1970 by John Ostrom. Currently located at the Teyler Museum in Haarlem, the Netherlands. It was the very first specimen, despite the classification error.
- Eichstätt Specimen (JM 2257): Discovered in 1951 or 1955 near Workerszell, Germany and described by Peter Wellnhofer in 1974. Currently located at the Jura Museum in Eichstätt, Germany. It is the smallest specimen and has the second best head. Possibly a separate genus, Jurapteryx recurva or species A. recurva.
- Solnhofen Specimen (BSP 1999): Discovered in the 1960s near Eichstätt, Germany and described in 1988 by Wellnhofer. Currently located at the Bürgermeister-Müller-Museum in Solnhofen. It was originally classified as a Compsognathus by an amateur collector. It is the largest specimen known and may belong to a separate genus and species, Wellnhoferia grandis.
- Munich Specimen (S6, formerly known as the Solnhofen-Aktien-Verein Specimen): Discovered in 1991 near Langenaltheim and described in 1993 by Wellnhofer. Currently located at the Paläontologische Museum München in Munich. What was initially believed to be a bony sternum turned out to be part of the coracoid (Wellnhofer & Tischlinger, 2004), but a cartilainous sternum may have been present. May be a new species, A. bavarica.
 The Munich Specimen
The Munich Specimen - Bürgermeister-Müller Specimen: A ninth, fragmentary specimen, was discovered in 1997. A further fragmentary fossil was found in 2004. It is kept at the Bürgermeister-Müller Museum.
- Thermopolis Specimen Discovered in Germany. Long in a private collection, described in 2005 by Mayr, Pohl, and Peters. Donated to the Wyoming Dinosaur Centre in Thermopolis, Wyoming, it has the best-preserved head and feet. The "Thermopolis" specimen, was described in the December 2, 2005 Science journal article as "A well-preserved Archaeopteryx specimen with theropod features", shows that the Archaeopteryx lacked a reversed toe—a universal feature of birds—limiting its ability to perch in trees and implying a terrestrial lifestyle. This has been interpreted as evidence of theropod ancestry. The specimen also has a hyperextendible second toe. "Until now, the feature was thought to belong only to the species' close relatives, the deinonychosaurs."



Synonyms
The synonymy of A. lithographica is extremely confused. Dozens of names have been published for the handful of specimens, most of which are simply spelling errors (lapsus). Originally, the name A. lithographica only referred to the single feather described by von Meyer. Swinton (1960) proposed that the name Archaeopteryx lithographica be officially transferred from the feather to the London specimen. The ICZN did suppress the plethora of alternative names initially proposed for the first skeleton specimens (ICZN, 1961), which mainly resulted from the acrimonious dispute between von Meyer and his opponent Johann Andreas Wagner (whose Griphosaurus - "enigmatic lizard" - was a vitriolic sneer at von Meyer's Archaeopteryx). In addition, descriptions of Archaeopteryx fossils as pterosaurs before their true nature was realized were also suppressed (ICZN, 1977).
If two names are given, the first denotes the original describer of the "species", the second the author on whom the given name combination is based. As always in zoological nomenclature, putting an author's name in parentheses denotes that the taxon was originally described in a different genus.
- Pterodactylus crassipes Meyer, 1857 [suppressed in favour of A. lithographica 1977 per ICZN Opinion 1070]
- Rhamphorhynchus crassipes (Meyer, 1857) (as Pterodactylus (Rhamphorhynchus) crassipes) [suppressed in favour of A. lithographica 1977 per ICZN Opinion 1070]
- Archaeopteryx lithographica Meyer, 1861 [nomen conservandum]
- Scaphognathus crassipes (Meyer, 1857) Wagner, 1861 [suppressed in favour of A. lithographica 1977 per ICZN Opinion 1070]
- Archaeopterix lithographica Anon., 1861 [lapsus]
- Griphosaurus problematicus Wagner, 1861 [nomen oblitum 1961 per ICZN Opinion 607]
- Griphornis longicaudatus Woodward, 1862 [nomen oblitum 1961 per ICZN Opinion 607]
- Griphosaurus longicaudatum (Woodward, 1862) [lapsus]
- Griphosaurus longicaudatus (Owen, 1862) [nomen oblitum 1961 per ICZN Opinion 607]
- Archaeopteryx macrura Owen, 1862 [nomen oblitum 1961 per ICZN Opinion 607]
- Archaeopterix macrura Owen, 1862 [lapsus]
- Archaeopterix macrurus Egerton, 1862 [lapsus]
- Archeopteryx macrurus Owen, 1863 [unjustified emendation]
- Archaeopteryx macroura Vogt, 1879 [lapsus]
- Archaeopteryx siemensii Dames, 1897
- Archaeopteryx siemensi Dames, 1897 [lapsus]
- Archaeornis siemensii (Dames, 1897) Petronievics, 1917
- Archaeopteryx oweni Petronievics, 1917 [nomen oblitum 1961 per ICZN Opinion 607]
- Gryphornis longicaudatus Lambrecht, 1933 [lapsus]
- Gryphosaurus problematicus Lambrecht, 1933 [lapsus]
- Archaeopteryx macrourus Owen, 1862 fide Lambrecht, 1933 [lapsus]
- Archaeornis siemensi (Dames, 1897) fide Lambrecht, 1933? [lapsus]
- Archeopteryx macrura Ostrom, 1970 [lapsus]
- Archaeopteryx crassipes (Meyer, 1857) Ostrom, 1972 [suppressed in favour of A. lithographica 1977 per ICZN Opinion 1070]
- Archaeopterix lithographica di Gregorio, 1984 [lapsus]
- Archaeopteryx recurva Howgate, 1984
- Jurapteryx recurva (Howgate, 1984) Howgate, 1985
- Archaeopteryx bavarica Wellnhofer, 1993
- Wellnhoferia grandis Elżanowski, 2001
The last 4 taxa may be valid genera and species.
"Archaeopteryx" vicensensis (Anon. fide Lambrecht, 1933) is a nomen nudum for what appears to be an undescribed pterosaur.